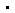Spindlin1蛋白识别组蛋白H3K4me3的结构示意图
10月17日,美国《国家科学院院刊》(Proceedings of the National Academy of Sciences)在线发表了生物大分子国家重点实验室许瑞明、饶子和课题组和北京生命科学研究所(NIBS)朱冰课题组合作的最新研究成果Distinct mode of methylated lysine-4 of histone H3 recognition by tandem tudor-like domains of Spindlin1。
组蛋白甲基化是表观遗传学的核心内容之一,其中关于甲基化的识别是近年来研究的热点。甲基化识别蛋白是表观遗传信号的执行者,它们特异地识别不同位点、不同形式的甲基化修饰,把信号传递给下游与其相互作用的分子,行使表观遗传的生物学功能。许瑞明课题组基于前期基础研究(Genes & Dev. 2003;Science 2006;Genes & Dev. 2009),此次在人源Spindlin1蛋白识别组蛋白H3第4位赖氨酸的三甲基化修饰(H3K4me3)研究中又取得了进一步成果。
人源Spindlin1蛋白最早作为纺锤体结合蛋白被发现(Dev. 1997)。2007年,饶子和课题组解析了Spindlin1蛋白的三维晶体结构(JBC 2007)。2011年,NIBS朱冰课题组发现Spindlin1定位在核内活性rDNA重复区,可识别组蛋白H3K4甲基化修饰,进而促进rDNA基因的表达(EMBO Reports 2011)。基于之前的研究结果,许瑞明课题组与朱冰研究员和饶子和教授开展了Spindlin1与组蛋白H3K4me3识别的结构机理研究。
结果表明,Spindlin1包含三个串联重复的Tudor结构域,其中只有Tudor II可以结合一个H3K4me3肽段(图A)。通过结构分析比较及体内外功能实验检测,研究人员们发现了Spindlin1蛋白识别H3K4me3的独特机制:首先,可结合甲基化赖氨酸残基的疏水口袋由4个芳香族残基组成(图B),比其他已知的Tudor结构域识别口袋都多了1个残基,这样的包围的更加紧密,同时对周围的环境要求也更加苛刻;其次,除了构成疏水口袋的芳香族残基,Spindlin1的极性天冬氨酸残基也与H3蛋白N端的精氨酸残基有多处较强的相互作用,分别对D184和D189残基进行突变,不同程度地影响了Spindlin1与H3K4me3肽段的结合能力,下调了体内rRNA基因的转录水平,这些极性相互作用确保了K4位点识别的特异性。
该项研究工作得到科技部、国家自然科学基金委员会和中国科学院的资助。(来源:中科院生物物理研究所)
特别声明:本文转载仅仅是出于传播信息的需要,并不意味着代表本网站观点或证实其内容的真实性;如其他媒体、网站或个人从本网站转载使用,须保留本网站注明的“来源”,并自负版权等法律责任;作者如果不希望被转载或者联系转载稿费等事宜,请与我们接洽。